
▪ Bible Physics ▪ Epinasty ▪ Plant Senescence Theory ▪ Scripture Evolution ▪ Socrates/Plato Civilization Cycle ▪ Speculations of Why Sexes Exist ▪
▪ 1986 Version ▪ 1995 Version ▪ 1999 Version ▪ 2003 Version ▪ 2007 Version a ▪ 2007 Version b ▪ Current Version ▪
Predictions
If the assumptions or postulates are true, I believe these must follow.
1. The reasons for the existence of the “Shoot” Growth Hormone Auxin include:
a. To attract all nutrients and Growth Hormones to new shoot cells that look like good investments for the plant in that they are producing high amounts of sugar and oxygen or most probably will do so in the future.
b. To induce orderly upward growth through apical dominance, so that the young, most efficient cells reach the most sunlight and the plant is “pointed” at the top and can pierce through the forest canopy if necessary. The Christmas tree growth pattern of apical dominance is also bottom heavy, adding to the balance and stability of the plant.
c. To cause the movement of sugar and oxygen down the phloem by attracting these nutrients from the leaves as Auxin is transported down the phloem.
d. To induce an increase in the minerals and water supply by:i) Causing more root cell division in concert with CK;
ii) Inducing new roots in the absence of CK; or
iii) Inducing a “jump start” to the water and mineral supply growth promoting ET synthesis (see below).
2. The reasons for the existence of the “Root” Growth Hormone CK similarly include:
a. To attract all nutrients and Growth Hormones to new root cells that look like good future investments to the plant, in that they are harvesting high amounts of water and minerals or most probably will do so in the future.
b. To induce orderly downward growth of the root through apical dominance, so that the young, most efficient root cells reach the deepest, most humid, soil, which is expected to be rich in minerals. The deeper the root goes, the more even the temperature the less energy is wasted on heating and cooling the root cells.
c. To cause water and minerals to move up the xylem, by attracting these nutrients from the roots into the transpiration stream.
d. To induce an increase in the sugar and oxygen supply by:i) Causing more shoot cell division in concert with Auxin;
ii) Inducing secondary bud outgrowth in the absence of Auxin; or
iii) Inducing a “jump start” in the sugar and oxygen supply growth by promoting GA/BR synthesis (see below).
3. The reasons for the existence of the Stress "Shoot" Hormone ET include:
a. The induction of hibernation of secondary buds under water and mineral shortage produced by Auxin and apical dominance. Because they are juvenile cells, ET may not be synthesized in large amounts there normally. Perhaps only enough ET is made to induce hibernation. As above, ABA is probably involved in slowing down the metabolism to a hibernating state. Low levels of ABA may be able to effect the hibernation alone. In fact, they may prevent senescence-inducing effects of ET.
b. To induce an increase in water and minerals by:i) Promoting the use of stored minerals and water;
ii) Inducing root hairs;
iii) Cutting back on some of the less efficient leaves that are unnecessary water and mineral sinks;
iv) Using the freed sugar, oxygen, water and minerals from the cannibalized shoots for new roots or better mature root performance;
vi) Inducing Auxin synthesis after a successful jump start of water and mineral level growth.c. To prune wounded and diseased tissue from the shoot, possibly in concert with ABA.
4. The reasons for the existence of the Stress "Root" Hormone GA/BR include:
 a. The induction of hibernation of secondary root buds under sugar and oxygen shortage produced by CK and root apical dominance. Because they are juvenile cells, perhaps GA/BR is not usually synthesized in large amounts there. Perhaps only enough GA/BR is made to induce hibernation. As above, ABA are probably involved in slowing down the metabolism to a hibernating state. Low levels of ABA may be able to effect the hibernation alone, and in fact may prevent the senescence-inducing effects of GA/BR.
a. The induction of hibernation of secondary root buds under sugar and oxygen shortage produced by CK and root apical dominance. Because they are juvenile cells, perhaps GA/BR is not usually synthesized in large amounts there. Perhaps only enough GA/BR is made to induce hibernation. As above, ABA are probably involved in slowing down the metabolism to a hibernating state. Low levels of ABA may be able to effect the hibernation alone, and in fact may prevent the senescence-inducing effects of GA/BR.
b. To induce an increase in sugar and oxygen by:
i) Promoting the use of stored sugar and oxygen;
ii) Inducing an increase in the efficiency of photosynthesis although in a more risky manner;
iii) Cutting back on some of the less efficient roots that are unnecessary sugar and oxygen sinks;
iv) Using the freed sugar, oxygen, water and minerals from the cannibalized roots for new leaves or better performance;
v) Inducing CK synthesis after a successful jump start of sugar and mineral level growth.c. To prune wounded and diseased tissue from the root, possibly in concert with ABA
5. The reasons for the existence of the “survival” hormones ABA include:
a. To slow down of metabolism of nutrient stressed cells so that their nutrient needs fall below those provided by the environment (they lower the “sustenance level” for the cell);
b. To induce senescence of nutrient stressed cells whose nutrient supply falls below the lowest possible sustenance level;
c. To slow down the metabolism of injured or diseased cells or tissue in order to limit damage;
d. To induce senescence of injured or diseased cells or tissue in order to limit the spread of the disease or of autocatalysis of tissue induced by ET.
e. To shut down the plant in case of an emergency. The most obvious condition like this is low water conditions.
6. The reasons for the existence of the “survival” hormone SA include:
a. To speed up the metabolism of healthy cells, so that all available nutrients are used up to the point of one limiting nutrient, to provide the most efficient growth or cell functioning;
b. To induce the division (in concert with Auxin and CK) of cells that have more than enough nutrients to support normal functioning and reach mature size;
c. To speed up the metabolism of healthy efficient cells near the site of injury or disease, to preserve them from autocatalysis of tissue induced by ET;
d. Possibly to re-induce cell division in tissue near cells that have senesced and abscised, in order to replace these cells.
e. To restart a plant into normal functioning after an emergency disappears. The most obvious of these conditions is the return of normal water conditions after rain.
7. In general, the following hormones inhibit senescence of the shoot and leaves: Auxin, CK, GA/BR and SA. ABA and ET are usually the only hormones that will inhibit shoot growth or induce shoot tissue senescence. In general, the following hormones inhibit the senescence of the root core and peripheral roots: Auxin, CK. ET and SA. ABA and GA/BR are normally the only hormones that inhibit root growth or initiate root senescence.
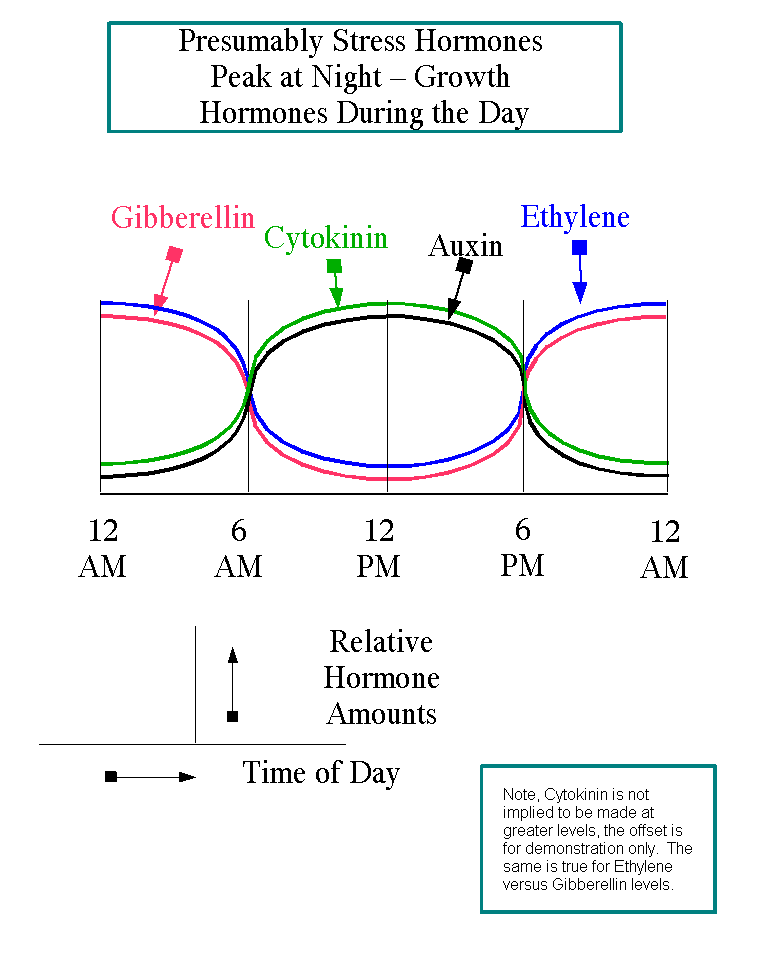
 8. Growth Hormones are at their highest levels during the day when there are the most opportunities for nutrient synthesis
8. Growth Hormones are at their highest levels during the day when there are the most opportunities for nutrient synthesis
and harvesting. Stress hormones reach their highest levels at night, when the plant must rely on nutrient reserves built up at night or prune inefficient tissue and use the resources to sustain itself through the period when little nutrient procurement is possible See the figure at below.
|
|
|
Click here for the full image. |
The plant can also slow down its metabolism at night and thus need fewer nutrients at night. This last strategy may be induced by ABA alone but is more likely to be induced by ET and GA/BR as well. The slower metabolism is also aided by the colder night temperatures.
9. Stress and ABA hormones are at their highest level at the beginning and end of the life of the plant or growing season, when a plant must rely on stored nutrients from senesced tissue and must be content with a slower rate of metabolism because of lower temperatures. Growth and SA hormones are generally at their highest level during the middle of the growth life of the plant or season.
10. Growth Hormones tend to move resources toward the edges of the plant to facilitate the procurement of more nutrients from the environment. This means they move resources to the top and bottom of the plant in the apical meristems but also perhaps move them laterally, to where the shoot and roots branch out like river deltas or capillary beds. Also, we can expect to find the highest concentration of the Growth Hormones at these edges of the plant, where the youngest cells reside. On the other hand, Stress Hormones tend to move resources inward, toward the stem and root core and closer to the soil line on both the root and shoot sides of the plant. The Stress Hormones engage the plant in a “hunkering down,” conservative, “smaller but stronger” posture. Also, we should expect to find the highest amounts of Stress Hormones near the soil line and in the shoot and root core.
11. It might seem that apical dominance could run away in growth like a cancer, but it doesn’t for seven reasons. These will be explained for Auxin and shoot apical meristems but a similar analysis could be done for CK and root apical dominance.
a. Auxin is transported down the stem through the phloem away from the shoot apex.
b. Auxin does induce xylem, so the shoot apical meristem hard wires a supply of water and minerals to itself. However, it also induces phloem, which takes the sugar and oxygen. away from the cells as they change to the mature cell morphology, so the multiplicative positive feedback effect of Auxin synthesis trails off as the meristem cells mature in the shoot.
c. Once a leaf has made xylem to itself as above, it only has to produce a small amount of Auxin to protect itself from senescence being initiated. The xylem guarantees that the leaf’s water and mineral supply cannot be hijacked by nearby meristematic tissue.
d. At any rate, as leaves increase in maturity and the plant grows, the older leaves move further and further away from the nutrient attractive forces of the Meristem (the leaves stay at the same level above the ground, and the meristem moves higher and higher off of it).
e. Relatively speaking also, the apical dominance may get weaker over time. As the older mature cells are closer to the source of the minerals and water, they are closer to the root and have “first dibs” on the supply of these nutrients. Meanwhile the apical meristem is moving further and further away from the source, the root.
f. If the apical meristem makes too much Auxin, this will induce ET both directly and indirectly. ET in turn inhibits the transport of Auxin and may inhibit its synthesis, thereby dampening the process. Auxin induces ET indirectly (and perhaps GA/BR and ABA) because the meristem’s draining of nutrients from surrounding tissue promotes the synthesis of these hormones. In addition to ET, we can expect at least ABA to inhibit Auxin synthesis.
g. There is an equally potent attractor of nutrients at the other end of the plant – the root apical meristem. These two “superpowers” probably work out a balance – an agreement to split the resources down the middle so to speak.
 12. A phenomenon called Epinasty (leaf stems – called petioles – lengthen and the leaf droops down and curls in on itself) is induced by ET. When ET is released it causes the senescence and abscission of some of the leaves. Since the leaves are the primary organs for harvesting oxygen, with the roots harvesting only some of the oxygen needed, the plant may be left with a rapidly developing partial anoxia. I hypothesize that the supply of oxygen becomes a growth and metabolism limiting factor more quickly than the supply of sugar. Thus, Epinasty is the plant’s choice as the lesser of two evils. The plant loses some photosynthesis efficiency because the leaves are no longer parallel to the ground with their face to the sun, but they may be able to trap more oxygen within their curled undersides. Alternatively or additionally, the Epinastic state may act like a sail, increasing the plant’s ability to trap oxygen and blowing up like a spinnaker. This may be bad physics, in that more gas diffusion occurs when the leaf surface is parallel to the direction of the wind. The leaves may also be trapping carbon dioxide, but this is unlikely since they only need to harvest enough carbon dioxide for their own photosynthesis needs. They do not support other cells with their supply of the gas. Thus, the leaves left after a bout of ET senescence need not configure themselves in any special way in order to get the carbon dioxide they need.
12. A phenomenon called Epinasty (leaf stems – called petioles – lengthen and the leaf droops down and curls in on itself) is induced by ET. When ET is released it causes the senescence and abscission of some of the leaves. Since the leaves are the primary organs for harvesting oxygen, with the roots harvesting only some of the oxygen needed, the plant may be left with a rapidly developing partial anoxia. I hypothesize that the supply of oxygen becomes a growth and metabolism limiting factor more quickly than the supply of sugar. Thus, Epinasty is the plant’s choice as the lesser of two evils. The plant loses some photosynthesis efficiency because the leaves are no longer parallel to the ground with their face to the sun, but they may be able to trap more oxygen within their curled undersides. Alternatively or additionally, the Epinastic state may act like a sail, increasing the plant’s ability to trap oxygen and blowing up like a spinnaker. This may be bad physics, in that more gas diffusion occurs when the leaf surface is parallel to the direction of the wind. The leaves may also be trapping carbon dioxide, but this is unlikely since they only need to harvest enough carbon dioxide for their own photosynthesis needs. They do not support other cells with their supply of the gas. Thus, the leaves left after a bout of ET senescence need not configure themselves in any special way in order to get the carbon dioxide they need.
12. In addition to the three behaviors high levels of Auxin can engender in the roots, there may be a fourth behavior. That is, the plant may try for quite a while to expand its minerals and water level, but if cell division, new root growth, and “jump starting” repeatedly prove unsuccessful, the plant may “decide” to store the extra sugar and oxygen until a more opportune time comes for the root. How the plant would measure repeated lack of success here, I don’t know.![]()
14. Finally, there is one other mysterious hormone effect that I would like to explain. It is again induced by ET. It is the phenomenon of the induction of adventitious roots (roots growing out of the shoot above the soil line and back into the ground) by the hormone under flooding conditions. I argue that this is because roots need to procure some oxygen from the soil, and flooding prevents this. With the roots starting out above the flooded soil in the atmosphere, they can absorb the oxygen needed and then dip back down into the soil to get the minerals and maybe even the water.